
What exactly is this phenomenon? We all learned that proteins are fundamental components of all living cells: our own, the bacteria that infect us, the plants and animals we eat. The hemoglobin that carries oxygen to our tissues, the insulin that signals our bodies to store excess sugar, the antibodies that fight infection, the actin and myosin that allow our muscles to contract, and the collagen that makes up our tendons and ligaments (and even much of our bones)—all are proteins.
To make proteins, `machines' known as ribosomes string together amino acids into long, linear chains. Like shoelaces, these chains loop about each other in a variety of ways (i.e., they fold). But, as with a shoelace, only one of these many ways allows the protein to function properly. Yet lack of function is not always the worst scenario. For just as a hopelessly knotted shoelace could be worse than one that won't stay tied, too much of a misfolded protein could be worse than too little of a normally folded one. This is because a misfolded protein can actually poison the cells around it.
The importance of protein folding has been recognized for many years. Almost a half-century ago, Linus Pauling discovered two quite simple, regular arrangements of amino acids—the a-helix and the b-sheet (see Fundamental Patterns of Protein Structure) - that are found in almost every protein. And in the early 1960s, Christian Anfinsen showed that the proteins actually tie themselves: If proteins become unfolded, they fold back into proper shape of their own accord; no shaper or folder is needed.
Of course, neither Pauling nor Anfinsen nor the committees that awarded them their respective Nobel prizes knew at the time that these discoveries would be so important for understanding Alzheimer's disease or cystic fibrosis. And when Pauling, at least, was doing his breakthrough studies, he could hardly have imagined the enormity of today's biotechnology industry. What scientists did know is that any process that was so fundamental to life as protein folding would have to be of the utmost practical importance.
But research did not stop with Pauling and Anfinsen. Indeed, we now know that Anfinsen's conclusions needed expansion: Sometimes a protein will fold into a wrong shape. And some proteins, aptly named chaperones, keep their target proteins from getting off the right folding path (see Molecular Chaperones). These two small but important additions to Anfinsen's theory hold the keys to protein folding diseases.
We've known since antiquity (but didn't know we knew) that protein folding can go wrong. When we boil an egg, the proteins in the white unfold. But when the egg cools, the proteins don't return to their original shapes. Instead, they form a solid, insoluble (but tasty) mass. This is misfolding. Similarly, biochemists have always cursed the tendency of some proteins to form the insoluble lumps in the bottom of their test tubes. We now know that these, too, were proteins folded into the wrong shapes.
Until recently, biochemists lacked the tools to study these insoluble lumps. Nor did they expect such masses would be particularly interesting. The prevailing view at the time was that the lumps were just hopelessly tangled and completely amorphous masses of protein fibers (aggregation). Researchers eventually discovered that these aggregates of incorrect folding could be highly structured, but before this crucial insight and before proper investigative tools were developed, biochemists simply threw their fouled test tubes away.
As far back as the start of this century, physicians have been noticing that certain diseases are characterized by extensive protein deposits in certain tissues. Most of these diseases are rare, but Alzheimer's is not. It was Alois Alzheimer himself who noted the presence of "neurofibrillary tangles and neuritic plaque" in certain regions of his patient's brain. Tangles are more or less common in diseases that feature extensive nerve cell death; plaque, however, is specific to Alzheimer's. The major question, which has only recently been answered, is whether plaque causes Alzheimer's or, like tangles, is a consequence of it.
Further investigation showed that neuritic plaque (unrelated to the plaque that clogs atherosclerotic blood vessels and causes heart attacks) is composed almost entirely of a single protein. Deposits of large amounts of a single, insoluble protein around the degenerating nerve cells of Alzheimer's disease eventually provided a key to understanding the disorder.
It was development of the biotechnology industry that unexpectedly spurred interest in insoluble protein gunk. This industry can produce proteins (often otherwise difficult-to-obtain human proteins) quickly and economically in bacteria. To their surprise, however, scientists who worked for biotech companies often found two things: protein that was supposed to be soluble instead precipitated as insoluble inclusion bodies within the bacteria and proteins that were supposed to be secreted into the surrounding medium instead got stuck at the bacterial cell wall.
This puzzling activity led scientists, almost for the first time, to seriously study just what goes wrong during protein folding.
In the decades after Anfinsen's work, the National Institutes of Health and the National Science Foundation continued to finance research in several laboratories. Working in relative obscurity, these protein biochemists tried to discover how a completely unfolded protein, with hundreds of millions of potential folded states to choose from, consistently found the correct one—and did so within seconds to minutes.
Could there be specific, critical intermediates (partially folded chains) in the folding process? This turned out to be a difficult question to answer. Partially folded chains don't stay that way very long; they become fully folded chains in a fraction of a second. Nevertheless, by the early 1980s researchers had not only found clear evidence for the existence of partially folded proteins, but also realized the key role these played in the folding process.
One study involved the difficulty in getting bovine growth hormone to fold properly. Although the unfolded proteins were not sticky, and the fully folded proteins were not sticky, the partially folded molecules stuck to each other—a first clue as to the origins of misfolded lumps (at least for purified proteins in test tubes). It still remained unclear why misfolding occurred in cells under certain circumstances but not under others.
The early 1980s also saw one of the first serious investigations of protein misfolding. These studies focused on temperature-sensitive mutations (mutations allowing growth at 75-F but not at 100-F) in the tailspike protein of bacteriophage P22. Neither bacteriophage P22, a virus that infects certain bacteria, nor its tailspike protein has any practical importance in themselves. Faced with thorny problems, however, scientists often look for experimental systems that will allow them to get a foothold or find a way around them. In this case, they thought that a large protein, whose folding passes through multiple stages, would be a good system for looking at folding pathways within cells. Many temperature-sensitive mutations had already been isolated in bacteriophages, but never examined for their effect on folding.
Their hopes were realized: The majority of the temperature-sensitive mutations they found, despite having only one amino acid altered, caused the tailspike protein to end up as insoluble gunk at high temperatures. Since these folding failures were occurring in bacterial cells that were growing in the laboratory, it was now possible to analyze what went wrong in a protein's folding process.
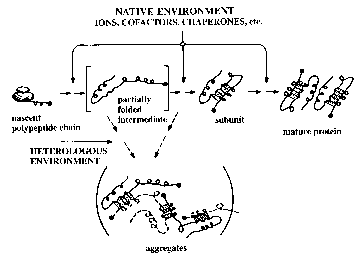 |
Partially folded intermediates at the junction between productive and off-pathway folding. Generalized pathways showing an inclusion body derived from an intermediate on the folding pathway. This illustration shows a speculative intermediate in the formation of an a/b protein in which a helical domain is docking against a sheet. In the inclusion body pathway, the same interaction proceeds between intermediates, resulting in a polymeric aggregate (3, 62). [Redrawn from FASEB J. 10, 58 (1997)] |
But this turned out not to be the case. If the mutant chains were allowed to fold up at low temperature, and were then heated, they were as stable as wild-type. It turned out to be a partially folded intermediate, on the route from the random shoelace to the correctly folded protein, that was sensitive to temperature. At higher temperatures these intermediates would stick to themselves and be unable to reach the properly folded state.
This turned out to be a general problem in the folding of many proteins: They have to pass through partially folded states in which they are delicately poised between folding all the way to the correct state or becoming seriously stuck as a result of premature entanglement with other molecules. Recognizing that it was the intermediates and not the fully folded protein that were in trouble opened the way to understanding some aspect of a range of diseases.
Over the past several years, collaborators have conducted similar studies in connection with a human disease. The minor differences between their results and others' are very revealing.
In the hereditary disease familial amyloidotic polyneuropathy (FAP), peripheral nerves and other organs are damaged by deposits of amyloid-type protein. Although the disorder is quite rare, extensive genetic studies have shown that the disease results from mutations in the protein transthyretin. As with the P22 tailspike protein, transthyretin contains large amounts of b-sheet structure and normally consists of several identical amino acid chains (four in this case) associated into a single, three-dimensional structure.
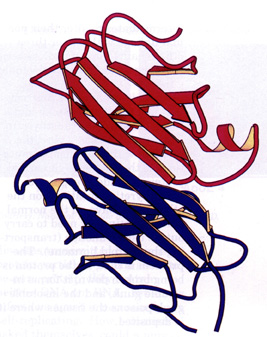 |
A `ribbon diagram' showing two molecules of the protein transthyretin docked together. The spiral coils of ribbon represent a-helix, while the flat arrows running alongside each other represent b-sheet. (Generated with Molscript by Scott Peterson, Texas A&M University) |
In both cases, however, the single-chain intermediates have structures that nature has designed for association with other chains of the same type. It apparently takes only a very small change in the shape of these intermediates to alter their normal linkage with two or three other chains into an endless series of linkages that creates insoluble gunk.
There is yet another contrast between the P22 tailspike mutations and those in transthyretin: From the P22 virus's view, the problem with the tailspike mutations is that not enough normal protein is made. People with transthyretin mutations, on the other hand, have all the normal transthyretin they need to carry out its usual function (transporting the thyroid hormone). The problem is that, as the protein is being broken down, it forms insoluble gunk, and the insoluble gunk poisons the tissues where it is deposited.
FAP is a rare disease; not so Alzheimer's, which afflicts 10 percent of those over 65 years old and perhaps half of those over 85. Every year Alzheimer's not only kills 100,000 Americans, but also costs society $82.7 billion to care for its victims.
In 1991, several different research groups found that individuals with specific mutations in their amyloid precursor protein developed Alzheimer's disease as early as age 40. The body processes amyloid precursor protein into a soluble peptide (small protein) known as Ab; under certain circumstances, Ab then aggregates into long filaments that cannot be cleared by the body's usual scavenger mechanisms. These aggregates then form the b-amyloid, which make up the neuritic plaque in Alzheimer patients. So the consistent association of amyloid precursor protein mutations with early-onset Alzheimer's has finally answered a long-debated question: the deposition of neuritic plaque is part of the pathway leading to the disease, not a late consequence of it.
To help understand the Ab aggregation process, researchers chemically synthesized fragments of the 40-amino-acid-long peptide. By using these fragments, they showed that the key step is getting started. Specifically, the precursor fragments have to form a specific nucleus, which then grows into the amyloid process. Possibly the slowness of this first step is why Alzheimer's disease is almost entirely limited to older people, and it could be that the mutations in amyloid precursor protein that lead to early-onset Alzheimer's are the ones that make it progress more quickly and easily.
Even so, Ab remains soluble in most people. Most individuals who develop Alzheimer's disease have the normal form of amyloid precursor protein, indistinguishable from that in people who never acquire the disorder. Why the same form of Ab aggregates in some people's brain but not in others' remains a mystery, although a recent discovery has suggested an intriguing possibility.
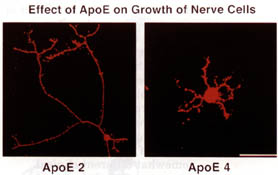
We know that people with different genetic variants of the protein apolipoprotein E (apoE) have quite different risks of developing Alzheimer's disease. Compared to those with the most common variant, known as apoE3, those with the apoE4 variant are significantly more likely to develop the disease. Some studies suggest that those with the apoE2 variant may be at lower risk, although other studies disagree.
These findings are particularly surprising because apoE is best known as part of the complex that transports cholesterol and other fatty materials in the bloodstream. What could a fat-transporting protein have to do with Alzheimer's disease? It may be significant that small amounts of this protein are associated with neuritic plaque and that apoE binds to Ab in the test tube. The results of this binding are in dispute, however.
Researchers report that adding apoE to a test-tube solution of soluble Ab causes rapid formation of plaque-type b-amyloid fibers—and that apoE4 does so more rapidly than apoE3. Others, however, have obtained opposite results: apoE prevents fibril formation. Thus, whereas some suggest that apoE acts as a pathological chaperone, one that actually promotes misfolding, other researchers believe that it exerts a normal chaperone's protective effect. In either case, apoE's influence on the folding of Ab may play a major role in development of Alzheimer's disease.
Perhaps the most interesting example of a protein folding disorder is Mad Cow disease and its human equivalent, Creutzfeldt-Jacob disease. These diseases, along with the sheep version known as scrapie, have had the scientific community in an uproar for years. They are infectious diseases transmitted by prions, or protein particles. Prions seem to be pure protein; they contain neither DNA nor RNA. Yet an infectious agent is necessarily self-replicating. How, scientists asked themselves, could a pure protein replicate itself?
 |
Courtesy: National Institute on Aging, Bethesda, MD. |
The protein whose aggregation damages nerve cells in Mad Cow disease is constantly being produced by the body. Normally, though, it folds properly, remains soluble, and is disposed of without problem. But suppose that somehow a small amount misfolds in a particular way so as to become a scrapie prion. If this scrapie prion bumps into a normal-folding intermediate, it shifts the folding process in the scrapie direction and the protein, despite its perfectly normal amino acid sequence, ends up as more scrapie prion. And the process continues: So long as the body keeps producing the normal protein, a little bit of scrapie prion can keep on creating more and then more. In effect, the prion is "replicating" itself without needing any nucleic acid of its own.
What old-school scientists find even more strange is that the process resembles something akin to genetics. Different strains of these diseases, with somewhat different clinical symptoms, `breed true' as they are transmitted from one animal or human to another. Moreover, these strain differences are associated with slight differences in the protein deposits that apparently cause the disease. (Scientists have recently used these strain differences to show that a few Britons truly have Mad Cow disease, the form seen in cattle, rather than the usual human form of Creutzfeldt-Jacob disease.)
Just as replication can occur without DNA or RNA, other experiments have shown how `genetics' is possible without nucleic acids. Thus, when researchers mix seed quantities of two different scrapie prion strains in separate test tubes with large amounts of normal protein, each test tube produces more of the specific scrapie prion strain that was added. That is, each strain induces the normal protein to fold in exactly the same way as the original seed. The strain breeds true in the test tube, just as it does in the body. Odd as it may seem, genetics without nucleic acid is truly possible in the world of protein folding.
Despite the examples of FAP, Alzheimer's disease, and Mad Cow disease, in which the problem derives from accumulation of toxic, insoluble gunk, many human diseases arise from protein misfolding leaving too little of the normal protein to do its job properly. The most common hereditary disease of this type is cystic fibrosis.
Recent research has clearly shown that the many, previously mysterious symptoms of this disorder all derive from lack of a protein that regulates the transport of the chloride ion across the cell membrane. More recently scientists have shown that by far the most common mutation underlying cystic fibrosis hinders the dissociation of the transport-regulator protein from one of its chaperones. Thus, the final steps in normal folding cannot occur, and normal amounts of active protein are not produced.
A hereditary form of emphysema shows an even greater analogy to the mutations studies in P22 tailspike protein. Investigators have found that one of the most common mutations producing this disorder greatly slows the normal folding process, just as the P22 temperature-sensitive mutations do. As with the tailspike mutations, the resulting buildup of a crucial folding intermediate leads to aggregation, which deprives affected individuals of enough circulating a1-antitrypsin to protect their lungs. Emphysema is the result.
As intriguing as these examples may be, there is a far more common instance of misfolding, which leaves too little normal protein to do its job. In this case, the protein's job is to block cancer development.
Over the past couple of decades, scientists have learned that most cancers result from mutation in the genes that regulate cell growth and cell division. The most common of these genes, involved in roughly 40% of all human cancers, is p53. The sole function of the p53 protein appears to prevent cells with damaged DNA from dividing before the damage is repaired (or to induce them to destroy themselves, if the damage cannot be fixed). In other words, p53 exists to prevent cells from becoming cancerous.
p53 mutations associated with cancer fall into two classes. The first keeps the protein from binding to DNA; the other makes the folded form of the protein less stable. In the second group, there is simply never enough properly folded protein around to block the division of DNA-damaged cells. It will be interesting to see how many of the p53 mutants fall into this second class and whether some way can be found to stabilize them.
The purpose of studying any human disease is to find ways to treat it. The story of protein folding has not yet led to treatments for the diseases involved, but this could happen within the next decade.
The key is to find a small molecule, a drug that can either stabilize the normally folded structure or disrupt the pathway that leads to a misfolded protein. Although many molecular biologists and protein chemists believe this will be quite difficult, others are more optimistic.
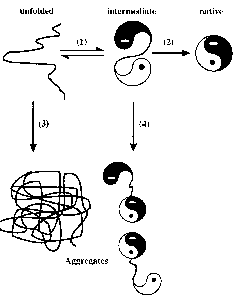 |
Folding and aggregation during protein renaturation. Correct folding reactions, leading to the native state [(1), (2)]. Irreversible aggregation reactions, starting from different conformations during the renaturation process [(3), (4)]. [From FASEB J. 10, 52 (1996)] |
Developing small-molecule therapies is quite straightforward for proteins like transthyretin that naturally bind small molecules, but these therapies are more difficult to apply to proteins that do not have a small-molecule binding site.
One of the few other groups currently publishing their research on small-molecule structure stabilizers is working to stabilize p53, an acknowledged `difficult target'. In fact, one laboratory has obtained encouraging results by using two different approaches.
Treatments based on our growing knowledge and contined research of protein folding are on the way. When they arrive, the saga that began with Pauling's fundamental studies of protein structure and Anfinsen's investigation of what some call `the second genetic code' will reach its practical fruition.
For a thorough discussion of protein folding, read a thematic issue on the topic in FASEB J. (1996) Protein Folding 10.
A very short review of protein folding with superb illustration is: Jonathan King (1993) The Unfolding Puzzle of Protein Folding, Technology Review, 58-61.
For an overview of the role of protein folding in human disease see Gary Taubes' (1996) Misfolding the Way to Disease, Science 271, 1493-1495.
For a short, comprehensive review of diseases due to protein misfolding, read Philip J. Thomas, Bao-He Qu, and Peter L. Pedersen (1995) Defective Protein Folding as a Basis of Human Disease, TIBS, 20, 456-459.
A good discussion of amyloid precursor protein and its role in Alzheimer's disease is: Celia Hooper (1991) An Exciting "If" in Alzheimer's, The Journal of NIH Research, 3 (April) 65-70.
For coverage of recent research on p53 and efforts to stabilize it, read: Rebecca L. Rawls (1997) Keeping Cancer in Check with p53, Chemical & Engineering News, 75 (February 18) 39- 41.
For everything you might want to know about prion disease, read Richard Rhodes (1997) Deadly Feasts, Simon & Schuster, New York, and a review by Neil Stahl and Stanley B. Prusiner (1991) Prions and prion proteins, FASEB J. 5, 2799-2807.
Return to FASEB Office of Public Affairs Homepage